


Dna ikileşmesi Nedir
Dna ikileşmesi, Replikasyon, Dna çoğalması ya da Dna sentezi, hücre bölünmesi öncesinde çift sarmallı Dna’nın kendini kopyalanması işlemidir.
Günümüzde yapılan araştırmalar sonucu, aynı tip hücrelerde Dna’nın kimyasal özelliğinin ve toplam mktarının nesilden nesile değişmeden aktarıldığı bilinmektedir. Buna göre Dna’nın tüm özellikleri aynı ata hücreden gelen benzer hücrelerde aynı kalmak zorundadır. Bu yüzden ister prokaryotik ister ökaryotik olsun her bir hücre mitoz bölünmeye hazırlanırken, Dna’lar kural olarak tüm uzunlukları boyunca bir ucundan diğer ucuna doğru kendilerini ikiler.
Watson ve Crick’in 1953’de yayımladıkları makaleleri, ikili sarmalın nasıl kendini eşleyeceği konusunda fikir vermektedir. “Yarı-saklı (semikonservatif) çoğaltma” olarak bilinen bu modelin geçerliliği o zamandan buyana değişmemiştir.
Çoğalmanın genel tarzı açıklık kazandıktan sonra araştırmalar, Dna sentezinin tüm ayrıntıları üzerine yoğunluk kazanmıştır. Günümüzde bilinen, Dna’nın kendini eşlemesi için sayısız enzim ve birçok proteine gerek duyduğudur. Sentez sırasındaki olayların karmaşıklığı bu araştırma alanının son derece aktif kalmasını sağlamıştır.
Dna kendini yarı-saklı eşlemeyle çoğaltır Watson ve Crick, sarmal açlıldığı takdirde, iki atasal zincir boyunca sıralanan bazların eşlenebilecekleri proteinleri kendilerine çekebileceklerini önermişlerdir. Buna göre, bazlar hidrojen bağlarıyla kendine uygun olan (örn. Adenin-Timinle, Guanin-Sitozinle) bazı çeker ve eşleşir. Her iki kalıp boyunca bu nükleotitler kovalent bağlarla polinükleotit oluşturdukça, birbiriyle özdeş iki Dna zinciri oluşacaktır. Kopyalanan her bir Dna molekülünde bir “eski” bir “yeni” zincir bulunacağından, bu tip bir çoğalma “yarı-saklı (semikonservatif) replikasyon” olarak tanımlanır.
Dna kopyalanması için, yine atasal zincirlerin kalıp olarak görev görmesine dayanan iki ayrı yol daha düşünülmüştür. Bunlar
Saklı (konservatif) replikasyon tamamlayıcı polinükleotit zincirleri yine aynı şekilde sentezlenir, ancak burada iki yeni zincir biraraya gelirken, atasal eski zincirler tekrar birleşir. Orjinal sarmal bu şeklide “korunur”.
Parçalı (dispersif) replikasyonda atasal zincirler kopyalama sırasında kırılır ve kırılan Dna parçaları iki yeni çift sarmal içinde dağılır. Böylece her bir zincirde hem yeni hem eski Dna bulunur. Üç olasılık içinde en karmaşık olan yol bu olduğu için, gerçekleşme ihtimali en zayıf olandır. Ancak, deneysel olarak teoride olması mümkündür. Her üç modelde baz eşlenikliğine dayandığı halde, yarı-saklı çoğalma en doğru olanıdır.
Tarihte
1958’de Meselson ve Stahl E.Coli ‘de yeni sentezlenen bir Dna’nın bir yeni bir de eski zincir içerdiğini göstererek yarı-saklı çoğalma konusundaki sorunu çözmüşlerdir. Taylor, Woods ve Hughes, baklanın kök uçlarıyla yaptıkları deneyde ökaryotlarda da yarı-saklı çoğalma olduğunu göstermişlerdir. Aynı dönemde Kornberg, E.Coli ‘den Dna Polimeraz I’i saflaştırmıştır. Kalıp ve öncü nükleozit trifosfatların bulunduğu ortamda, bu enzimin in vitro Dna sentezi yapabileceğini göstermiştir. Daha sonra Dna Polimeraz II ve III izole edilmiş ve polimeraz III, in vivo Dna kopyalanmasından sorumlu enzim olarak tanınmıştır.
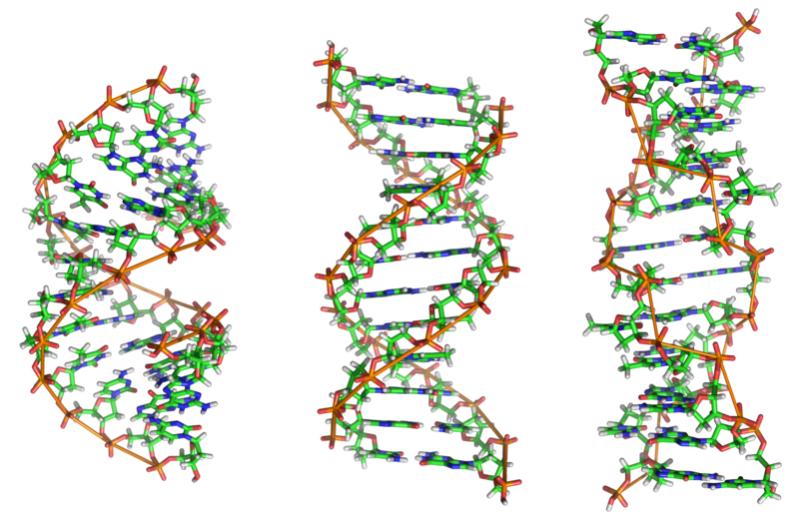
Dnanın Yapısı
Dna, tüm hücrelerde bulunan, nesilden nesile aktarılabilen çift bir moleküldür. Bu çift molekül, bir sarmaşığın dalları gibi birbiri çevresinde dönerek bir sarmal oluşturur. Sarmaşık dalına benzer her molekül, bir Dna “ipliği”dir. Bu iplikler birbirlerine kimyasal olarak bağlanmış nükleotitlerden oluşur. Nükleotitler ise bir şeker, bir fosfat ve bir de dört çeşit azotlu bazlardan birisinden oluşur. Bu dört çeşit baz, Adenin, Timin, Sitozin ve Guanindir. Sırası ile A, T, C ve G harfleri ile kısaltılırlar. Her baz diğer bazların yalnızca bir çeşidi ile hidrojen bağları kurabilir, kural olarak A ile T, C ile ise G bağ kurabilir.
Dna Replikasyonu
Dna molekülünün ikileşmesinde, sarmalın kollarnı birbirine bağlayan zayıf hidrojen bağları fermuar gibi açılır her iki kolda, eşlerinden ayrılan pürin ve pirimidin uçlarını açıkta bırakır. Hücrenin sitoplazmasında bulunan çeşitli nükleotitlerin iki kol açıldıkça, kollarda bulunan uygun bazların karşılarına gelmeleriyle kendini eşleme başlamış olur. Dna’nın ikili sarmalı birbirinden ayrıldığı zaman, kural olarak Adenin grubu Timin grubuyla, Guanin grubuysa Sitozin grbuyla birleşerek yerlerini alırlar. Diğerleri uymadıkları için geri çevrilirler. Yine aynı şekilde, eski zincirdeki Adeninler Timinlerle, Sitozinler Guanin gruplarıyla ikili sırayı tamamlamak için birleşirler. Bütün nükleotitler eşlendiğinde ise, yeni zincir oluşturulmuş, Dna kendini eşlemiştir. Kopyalanan yeni Dna iplikleri tamamen aynıdır, ancak nadiren çoğalmadaki hatalar nedeniyle kopyalama mükemmel olmaz (bkz. mutasyon).
ikileşme Orijini ve Çatalı
Kromozom üzerinde replikasyonun başladığı bölge “replikasyon orjini” olarak adlandırılır. Kromozom üzerinde replikasyonun olduğu noktada sarmala ait zincirlerin açılmasıyla meydana gelen çatala “replikasyon çatalı” denir. Bu çatal, önce sentezin orjin noktasında meydana gelir ve replikasyon devam ettikçe ilerler. Replikasyon çift yönlü ise, orjinden itibaren zıt yöne doğru ilerleyen iki replikasyon çatalı oluşur. Replikasyonun orjini ve yönü ile ilgili kanıtlar açıktır.
Prokaryotlarda Dna ikileşmesi
Dna replikasyonunda, ikili sarmal açılır ve sentezin başladığı yer olan replikasyon çatalı oluşur. Proteinler açılan sarmalı kararlı kılar ve replikasyon çatalının önünde oluşan sarılma gerilimini hafifletirler. Sentez, kalıp boyunca belirli bölgelerden RNA Primazın, Dna Polimeraz III’ün polimerizasyonu başlatabileceği serbest 3′-OH ucunu sağlayan kısa bir RNA parçasını sentezlemesiyle başlar. ikili sarmalın antiparalel yapısından dolayı polimeraz III, kesintili zincirde 5′-3’ yönünde sürekli Dna sentezi yapar. Kesintili zincir denen karşı zincirde kısa Okazaki fragmanları sentezlenir ve bu fragmanlar daha sonra Dna Ligaz ile birleştirilir. Dna Polimeraz I, RNA primerini uzaklaştırır ve yerine Dna sentezler, ortaya çıkan polinükleotidler (Dna parçaları) Dna Ligaz ile birleştirilir. Dna replikasyonunda yer alan birçok molekülü etkileyen pekçok mutant bakteri ve faj genlerinin izole edilmesi, tüm replikasyon işleminin karmaşık genetik kontrolünün aydınlanmasına yardımcı olmuştur.
Replikon, oriC ve ter
Cairns, izotoplar kullanarak, otoradyografi yöntemiyle replikasyonu izlemiş ve E.Coli’de replikasyonun tek bir noktadan (orjinden) başladığını göstermiştir. Bu özgül bölgeye oriC denilmiştir. Bu bölgenin konumu E.Coli üzerinde haritalanmış ve 245 baz içerdiği saptanmştır. Bu konuda yapılan başka araştırmalarda da, replikasyonun iki yönlü olduğu ve oriC’nin her iki yönünde hareket ettiği gösterilmiştir. Bu durumda replikasyon ilerledikçe ayrı yönlere doğru birbirinden uzaklaşan iki replikasyon çatalı oluşturur. Bu çatallar tüm kromozom yarı-saklı eşleştikten sonra, “ter” olarak adlandırılan sonlanma bölgesinde birbiriyle birleşir. Bir orjinden replikasyon başladıktan sonra eşleşen Dna’nın uzunluğunun bir birim olduğunu belirten terim “replikon”dur. Buna göre, bakteriyofaj ve bakterilerde Dna sentezi bir noktadan (oriC), başlayıp bir noktada (ter) biter. Bakteriler, tek ve büyük bir kromozoma sahip oldukları için, kromozomun tümü bir “replikon”dur.
Dna Polimeraz I, II ve III
Replikasyonun yarı-saklı ve iki yönlü olduğu anlaşıldıktan sonra, birçok moleküler çalışma Dna kalıbı üzerinden tamamlayıcı uzun polinükleotit zincirlerinin gerçek sentezinin nasıl olduğunu anlamaya yönelmiştir. Bu çalışmalarda kullanılan mikroorganizmalarda, sentezde gerekli olan, Dna Polimeraz I, II ve III olarak bilinen enzimlerin varlığı görülmüştür.
Ökaryotlarda Dna ikileşmesi J.H. Taylor, P.Woods ve W.Hughes 1957’de ökaryotlarda da replikasyonun yarı-saklı olduğunu gösteren kanıtı sunmuşlardır. Vicia faba (bakla) bitkisinin kök uçlarıyla yaptıkları deneyde Dna’yı 3H-timidin ile işaretleyip, otoradyografisini çekmişler ve replikasyonu izlemeyi başarmışlardır. Buradaki replikasyonun yarı-saklı olduğunu kanıtlamışlardır.
Ökaryotlardaki Dna replikasyonu prokaryotlardakine benzer ancak daha karmaşıktır. Her iki sistemde de Dna ikili sarmalı “replikasyon orjini”nden açılarak iki “replikasyon çatalı” meydana gelir. Dna polimerazın yönlendirdiği sentez, kesintisiz zincirde ve kesintili zincirde çift yönlü olarak devam eder. Prokaryotlardan en önemli fark olarak, ökaryotlada birçok “replikasyon orjini” ve sentezi yönlendiren daha farklı Dna Polimerazlar bulunmasıdır. Bunun nedenleri şöyle açıklanır:
Ökaryotlarda, prokaryotlara göre daha fazla gen vardır Ökaryotik polimerazın saniyede 50 nükleotit olan okuma hızı, prokaryotik polimeraza göre 20 kat yavaştır Çoklu replikasyon orjini ile ilk bulguların çoğu bir maya olan Saccharomyces cerevisiae’den elde edilmiştir. Mayadan elde edilen bu repliksyon orjinlerine “özerk replike olan diziler” (ARS) denir. Hücre döngüsünün G1 fazı sırasında bütün ARS dizilerine bazı protein grupları bağlanır ve “orjin tanıma kompleksi” (ORC) meydana gelir. Bu tanıma kompleksleri G1 fazında oluştuğu ve S fazından önce sentez başlamadığı için, sentezin gerçek başlama sinyalinde yer alan daha başka pronteinler de bulunmaktadır. Bu proteinlerin en önemlileri özgül kinazlardır. Kinazlar, hücre döngüsünün ayrılmaz bir parçası olan fosforilasyonun kilit enzimleridir. Kinazlar, ORC’ye bağlandıklarında, Dna polimerazın bağlanmasına açık olan bir “ön tanıma kompleksi” (pre-RC) oluşur. pre-RC’ye bağlanacak Dna polimerazlar, ökaryotik replikasyonun en karmaşık yönüdür. Buna göre, 6 farklı tipte Dna polimeraz formu saflaştırılıp, çalışılmıştır:
Ökaryotlarda doğrusal kromozom uçlarının (telomerler) replikasyonda ortaya çıkan özel sorun, RNA içeren özgün bir enzim olan telomeraz enzimiyle çözülür.
Genetik moleküller arasındaki rekombinasyon, Dnalglfrglelglergleplgperogpeorogrep zincirlerini kesen, tekrar sıraya koyan ve tekrar birleştiren bir dizi enzim varlığına dayanır. Gen dönüşümü olayı, bu değiş-tokuşlar sırasında yanlış eşleşme onarımı ile gerçekleştirilen sentez ile en iyi şekilde açıklanabilir